Results and Discussion
Of all the models in this website, I have examined this Public Goods model most thoroughly to date. You can find an in-depth analysis in "Altruism pays off in group-structured populations through probable reciprocity" (PDF on bioRxiv). Here is a short excerpt from it.
Selection only: population development without reproduction and migration
We start with the examination of pure selection events without reproduction, migration and mutation. So it is only about the elimination of individuals, whether their achieved scores reach the threshold for survival or not. Used values are costs K = 5, benefit B = 10 and threshold T = 3.
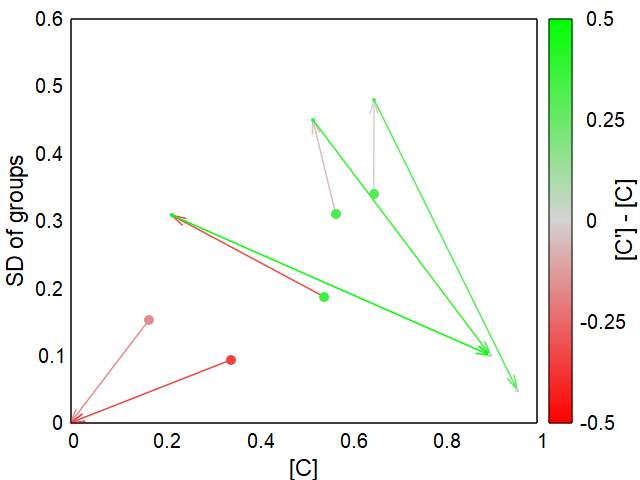
From the initial population, a maximum of two steps lead to a fixed final state either with a high proportion of C ([C]) or without any C, which is equivalent to the extinction of the population, since D alone cannot survive under the conditions mentioned.
The three greenish starting populations in Fig. 1 achieve a gain in fitness up to the final population, i. e. a relative increase in C. In one case there is initially a marked decrease in fitness, but in the final state all three are positive. A second component is noticeable in all three cases: the sometimes significant increase in variance after the first iteration.
The change in fitness is only half the battle. Just as important is the change in variance (SD of groups) in order to be able to describe the development of a population. That would be going too far here, but you can find more information on this in my paper, see above.
The results can be summarized as follows: Variation among groups is crucial for the persistence of altruistic traits in the population.
On the threshold of altruism
So far, we have only considered threshold models with costs K = 5, benefits B = 10 and threshold T = 3, and we have seen that altruism can persist as soon as the population is divided into groups. Let us now look at the development of populations with other costs, benefit values and thresholds. The interesting case here is when the benefit for the group is greater than the cost for C, that is B > K.We see that stable altruism is only possible if the threshold lies between 0, that is a pure D population ([C]=0), and B – K, which corresponds to a pure C population ([C]=1).
| Threshold T | Effect | Dominance |
|---|---|---|
| T ≤ -K | No selection pressure on C and D, therefore starting population already freezes. | C≈D |
| -K < T ≤ 0 | Some selection pressure only on C, as only C can have scores < 0. Below-threshold C are eliminated until groups become fixed. | D>C |
| 0 < T ≤ B – K | Selection pressure on C and D, but C are relatively fitter than D. Range stable altruism! | C |
| B – K < T | Lethal selection pressure on C and D. Extinction of the population. | † |
Standard model: Population dynamics with reproduction, migration, mutation and additional mortality
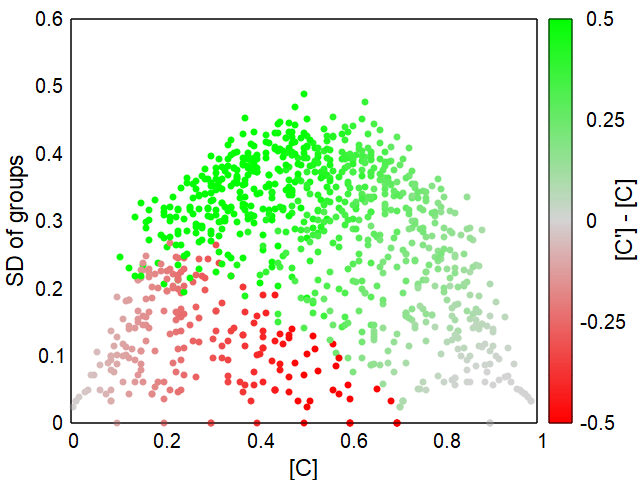
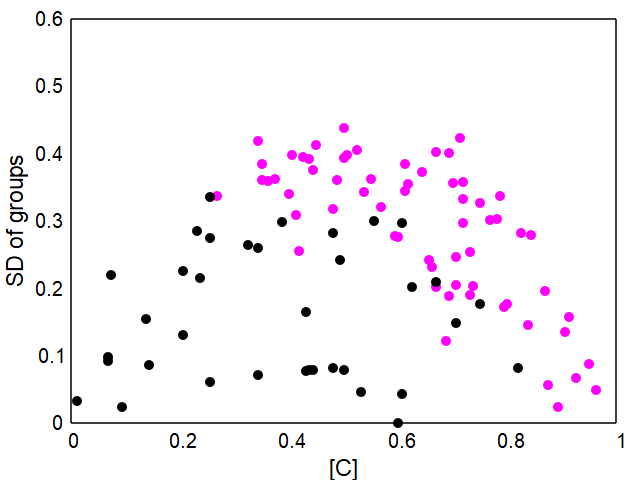
Adding reproduction, migration between groups and additional mortality factors to the model results in more dynamic systems.The standard model uses again a initial population of 16 groups of 10 individuals each, whether of type C or D is again chosen at random. The other parameters can be seen in the simulation interface after clicking the "standard model" example. About 64% of the randomly set initial populations develop stable populations of mainly altruists (C) as shown in Fig. 3 and 4.
Details and the parameter ranges that lead to altruistic populations can be found in my paper mentioned above (or open it here).
Evolution of altruism
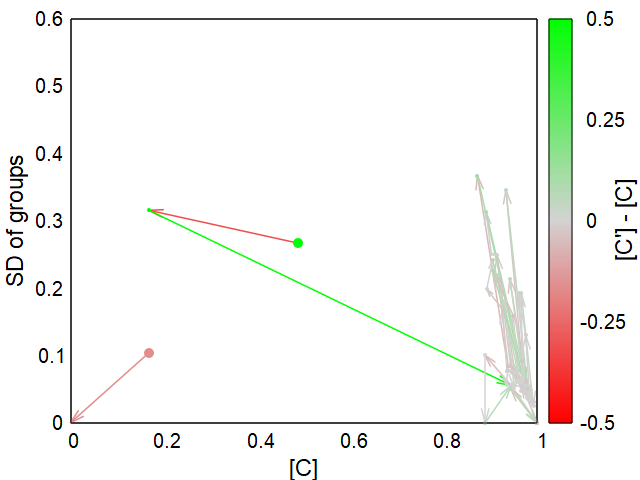
As described above, starting populations with predominantly altruists are a prerequisite for stable altruistic populations. Nevertheless, I found two scenarios in which an altruistic trait can be established in a purely selfish starting population.Evolve #1
In the first approach, there is initially no selection pressure on C and D, i.e. the “altruistic” trait is neutral, or in the context of the model when T ≤ -K (or T ≤ 0 if K<B/N). If there is only little migration between the groups, this automatically results in large variance between the groups, because almost pure C and D groups are created by chance and inheritance. This gives a favorable starting situation for altruism: high variance between groups. If the selection pressure now increases so that the threshold lies in the range 0 < T ≤ B - K, a stable population with predominantly altruists quickly develops due to the G-effect.Evolve #2
For the second approach, it is crucial that the sub-threshold mortality rate is less than 100% and here, too, restricted migration is important. It works best when empty groups are filled by migration and there is otherwise little exchange between existing groups. If the sub-threshold mortality is low, the altruistic trait cannot establish even if threshold level would allow this (0 < T ≤ B – K). Above a certain sub-threshold mortality rate, however, the altruistic trait suddenly establishes itself and then remains even if the sub-threshold mortality rate is increased to 100%.Of crucial importance for the evolution of altruism in both scenarios is limited migration between the groups and an increasing selection pressure for all individuals, whether by shifting either the threshold or the sub-threshold mortality.
References and further reading
Ackermann, M., Stecher, B., Freed, N.E., Songhet, P., Hardt, W.-D., Doebeli, M., 2008. Self-destructive cooperation mediated by phenotypic noise. Nature 454, 987–990. https://doi.org/10.1038/nature07067
Bin Masroni, M.S., Koay, E.S.-C., Lee, V.K.M., Ng, S.B., Tan, S.Y., Tan, K.M., Archetti, M., Leong, S.M., 2025. Sociobiology meets oncology: unraveling altruistic cooperation in cancer cells and its implications. Exp Mol Med 57, 30–40. https://doi.org/10.1038/s12276-024-01387-9
Capp, J., Thomas, F., Marusyk, A., M. Dujon, A., Tissot, S., Gatenby, R., Roche, B., Ujvari, B., DeGregori, J., Brown, J.S., Nedelcu, A.M., 2023. The paradox of cooperation among selfish cancer cells. Evolutionary Applications 16, 1239–1256. https://doi.org/10.1111/eva.13571
Coggan, H., Page, K.M., 2022. The role of evolutionary game theory in spatial and non-spatial models of the survival of cooperation in cancer: a review. J. R. Soc. Interface. 19, 20220346. https://doi.org/10.1098/rsif.2022.0346
Cremer, J., Melbinger, A., Wienand, K., Henriquez, T., Jung, H., Frey, E., 2019. Cooperation in Microbial Populations: Theory and Experimental Model Systems. Journal of Molecular Biology 431, 4599–4644. https://doi.org/10.1016/j.jmb.2019.09.023
Darwin, C., 1888. The descent of man: and selection in relation to sex. John Murray, Albemarle Street.
Dugatkin, L.A., 2017. The evolution of altruism. Vestn. VOGiS 21, 487–491. https://doi.org/10.18699/VJ17.267
Eshel, I., 1972. On the neighbor effect and the evolution of altruistic traits. Theoretical Population Biology 3, 258–277. https://doi.org/10.1016/0040-5809(72)90003-2
Fletcher, J.A., Doebeli, M., 2009. A simple and general explanation for the evolution of altruism. Proc. R. Soc. B. 276, 13–19. https://doi.org/10.1098/rspb.2008.0829
Fletcher, J.A., Zwick, M., 2004. Strong altruism can evolve in randomly formed groups. Journal of Theoretical Biology 228, 303–313. https://doi.org/10.1016/j.jtbi.2004.01.004
Hamilton, W.D., 1964. The genetical evolution of social behaviour. I. Journal of Theoretical Biology 7, 1–16. https://doi.org/10.1016/0022-5193(64)90038-4
Hardin, G., The Tragedy of the Commons. Science162,1243-1248(1968). DOI:10.1126/science.162.3859.1243
Killingback, T., Bieri, J., Flatt, T., 2006. Evolution in group-structured populations can resolve the tragedy of the commons. Proc. R. Soc. B. 273, 1477–1481. https://doi.org/10.1098/rspb.2006.3476
Kropotkin, P., 1902. Mutual aid: A factor of evolution. McClure Phillips & Co.
Maynard Smith, J., Price, G. The Logic of Animal Conflict. Nature 246, 15–18 (1973). https://doi.org/10.1038/246015a0
Nowak, M.A., 2012. Evolving cooperation. J Theor Biol 299, 1–8. https://doi.org/10.1016/j.jtbi.2012.01.014
Nowak, M. A. & May, R. M., 1992. Evolutionary games and spatial chaos. Nature 359, 826–829. (doi:10.1038/ 359826a0)
Nowak, M. A., Tarnita, C. E., Antal T., 2010. Evolutionary dynamics in structured populations. Phil. Trans. R. Soc. B. 365, 19–30 (doi:10.1098/rstb.2009.0215)
Okasha, S., 2020. Biological Altruism, in: Zalta, E.N. (Ed.), The Stanford Encyclopedia of Philosophy. Metaphysics Research Lab, Stanford University.
Pepper, J.W., 2000. Relatedness in Trait Group Models of Social Evolution. Journal of Theoretical Biology 206, 355–368. https://doi.org/10.1006/jtbi.2000.2132
Price, G.R., 1970. Selection and Covariance. Nature 227, 520–521. https://doi.org/10.1038/227520a0
Sigmund, K., Hauert, C., 2002. Altruism. Current Biology 12, R270–R272. https://doi.org/10.1016/S0960-9822(02)00797-2
Sober, E., Wilson, D.S., 1998. Unto Others: The Evolution and Psychology of Unselfish Behavior, Emersion: Emergent Village Resources for Communities of Faith Series. Harvard University Press.
Steiner, K.F., 2021. The Good, the Bad and the Stochastic: How Living in Groups Innately Supports Cooperation. bioRxiv 2021.02.21.431661; doi: https://doi.org/10.1101/2021.02.21.431661
Steiner, K.F., 2024. Altruism pays off in group-structured populations through probable reciprocity. bioRxiv 2024.01.20.575560. doi: https://doi.org/10.1101/2024.01.20.575560
Trivers, R.L., 1971. The Evolution of Reciprocal Altruism. The Quarterly Review of Biology 46, 35–57. https://doi.org/10.1086/406755
Wilensky, U., 1999. NetLogo. http://ccl.northwestern.edu/netlogo/. Center for Connected Learning and Computer-Based Modeling, Northwestern University. Evanston, IL.
Wilson, D.S., 1975. A theory of group selection. Proc. Natl. Acad. Sci. U.S.A. 72, 143–146. https://doi.org/10.1073/pnas.72.1.143
Wilson, D.S., Wilson, E.O., 2008. Evolution “for the Good of the Group.” American Scientist 96, 380–389.